
J.D. van der Toorn (1997,1998)
A survival guide to survival rates
Updated October 1999
The survival of marine mammals in captivity is often the subject of heated discussions. Interestingly, these discussions usually focus on cetaceans. The discussions are often complicated by a general lack of understanding of the subject matter. This can result in incorrect representation of the available data and comparisons of unrelated parameters. The terminology involved is not straightforward and can be confusing (Fad, 1996).
In this paper, I will discuss the terminology involved, the calculations that must be done to derive survival rates and life expectancies and I will look at the presentation of the data and how that can influence the message. A few examples will be given.
First, let's have a look at some of the terms. All these terms are derived from population dynamics modelling.
Note that all these parameters are calculated for non-calf or non-pup animals (age >
1 year) (Small and DeMaster, 1995). The survival of calves and pups is incorporated
in the calculation of reproductive rates. The focus of this paper is on survival rates.
The reproductive parameters will be discussed only briefly.
Some background on reproductive ratesThe following terms can be encountered in discussions about reproduction, as mentioned in a) Olesiuk et al (1990) and b) Wells and Scott (1990):
|
In the following calculations, it is assumed that the ASR and DSR are independent of the actual age of the animals. This is probably not completely accurate. It is likely, that the survival rates differ for different age classes (Barlow and Boveng, 1991). However, the data available to date is insufficient to reliably determine age class dependent survival rates for most marine mammal species (due to a limited sample size) (Small and DeMaster, 1995).
The DSR can be calculated as follows (DeMaster and Drevenak, 1988):
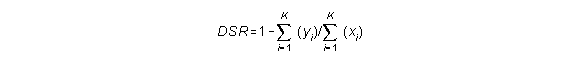
where:
Or, in a simpler form (Small and DeMaster, 1995):
![]()
The annual survival rate ASR can be derived from the Daily Survival Rate DSR by
raising the DSR to the power 365.25 (the average number of days in a year), as
follows:
![]()
The annual mortality rate AMR is, by definition:
![]()
If we want to estimate the life expectancy for the population (the average number of
years survived), we first need to calculate the total animal years. If we start out with
N0 animals, we need to know how many of them are left after one year, two years,
etc. After one year, we have:
![]()
In this and the following equations s is the ASR. After two years:
![]()
And after n years:
![]()
If we add them all together, we get:
![]()
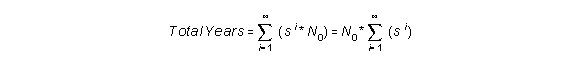
If we take infinitely small steps, we can rewrite this formula to:
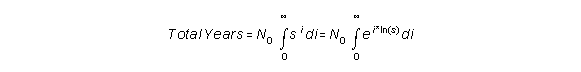
This results in:
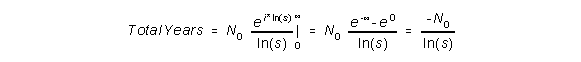
To get the life expectancy, which is the average number of years, we need to divide
the total number of years by the number of animals in the sample:
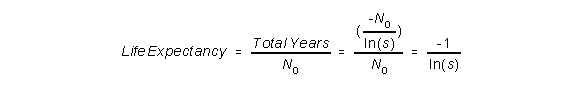
If we play around a bit with this formula, we can see that the life expectancy is very
sensitive to errors. If there is only a small error in the estimate of the ASR, this can
result in an error of several years in the derived estimate for the life expectancy. For
that reason, life expectancy should not be used when comparing populations,
captive or wild (DeMaster and Drevenak, 1988). See the table below for some survival rates,
the corresponding life expectancies and the increase in both compared to the previous value in
the table. You'll see that a 1% increase in survival rate results in a huge increase in
life expectancy estimate for higher ASR values. If you want to play around with some numbers yourself,
go to the ASR to Life Expectancy Converter on the Survival Rates Toolkit page.
| ASR | ASR increase | Life expectancy | LE increase |
| 0.920 | 11.99 | ||
| 0.930 | 1.09 % | 13.78 | 7.26 % |
| 0.940 | 1.08 % | 16.16 | 17.27 % |
| 0.950 | 1.06 % | 19.50 | 20.67 % |
| 0.960 | 1.05 % | 24.50 | 25.64 % |
| 0.970 | 1.04 % | 32.83 | 34.00 % |
| 0.980 | 1.03 % | 49.50 | 50.78 % |
The reproductive ratesThe neonate survival is the complement of neonate mortality:SRn = 1 - MRn From this we can calculate the relation between fecundity rate and fertility rate and between birth rate and recruitment rate: FEC = FER * SRn = FER * ( 1 - MRn ) REC = BR * SRn = BR * ( 1 - MRn ) Where:
|
So far, we have not yet mentioned the longevity or maximum age. The reason for this is that there is no direct correlation between survival rate and longevity. Longevity is basically an incidental finding: it is determined by the oldest animal you find in your sample. You can only determine the longevity of a group of animals after all the members of that group have died (Duffield and Wells, 1991).
Sometimes, average age is used in comparing wild and captive populations (Duffield and Wells, 1991). The average age is calculated by adding the ages of all the animals in the population and dividing this by the number of animals. This is however of a very limited use. Such a comparison will only yield useful data if both populations have been stable for a long period of time (no increase or decline) (Barlow and Boveng, 1991). Only in a stable population, the calculation of the average age will yield the same result as the calculation of the life expectancy. If the size of a population is changing however, the average age can shift either way.
Let's consider a declining population. There can be a number of reasons for the decline, such as:
Also increases in population size can shift the average age either way:
So, a higher average age does not necessarily mean that a certain population is
doing better than another population with a lower average age.
 Next page
Next page |
|
 Back to the Online Papers Page
Back to the Online Papers Page |
 Back to the Publications Page
Back to the Publications Page |